| NIES-MCC | KU-MACC | Tree to Strain | Japanese | English |
| 生物 / 真核生物 | |
|
オピストコンタ(後生動物、襟鞭毛虫、菌類 など) |
|
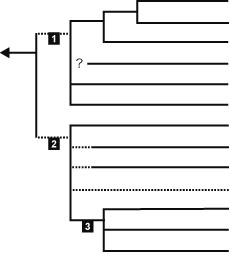
※点線は非単系統である可能性があるもの 1. ユニコンタ Unikonta 2. バイコンタ Bikonta (= Plantae sensu Nozaki et al. 2007) 3. ハロサ亜界 Harosa (= SAR clade) |
|
| Refereces |
|
|
我々ヒトを含む後生動物や陸上植物など多細胞生物のほとんどは真核生物であり、酵母や繊毛虫、アメーバ、ミドリムシなど単細胞性の真核生物も多い。真核生物では、遺伝情報であるDNAが線状で多数のタンパク質とともにヌクレオソーム/染色体という複合体を形成している。染色体は核とよばれる二重膜(核膜)で区切られた空間に局在する。このため転写と翻訳の場が空間的に区画化されている。おそらく全ての真核生物は真正細菌 (α-プロテオバクテリアに近縁な生物) の細胞内共生に起源をもつミトコンドリア(またはそれと相同な構造)をもっており、また小胞体やゴルジ体などの内膜系が発達している点でも原核生物とは異なる。真核生物は基本的に従属栄養性(捕食・吸収)であるが、植物 (灰色植物、紅色植物、緑色植物) の共通祖先がシアノバクテリアとの細胞内共生によって葉緑体を獲得し、光独立栄養が可能となった。その後、この葉緑体は二次共生、三次共生によってさまざまな真核生物に転移している (不等毛植物など)。 分子系統学的研究の発展から、近年では真核生物の中にいくつかのスーパーグループ(オピストコンタ、アメーボゾア、リザリア、アルベオラータなど)が存在していることが判明してきている。さらに遺伝子融合などの情報から、真核生物はユニコンタ(オピストコンタ + アメーボゾア)とバイコンタ(植物などそれ以外の真核生物)に分かれることが提唱されているが、その妥当性はいまだはっきりしていない。 |
 1: Vanella (アメーボゾア). 2: 有孔虫 (リザリア). 3: Cryptomonas (ハクロビア). 4: アミメネコゼミジンコ (オピストコンタ). 5: カヤモノリ (ストラメノパイラ). 6: コモチマンネングサ (植物). 7: ゾウリムシ (アルベオラータ). 8: Phacus (エクスカヴァータ). |